
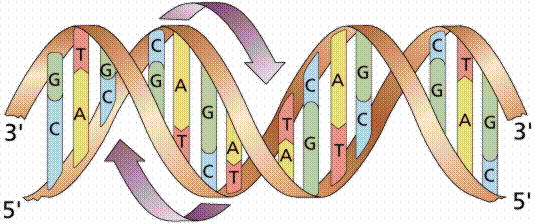
 En
los años 20, el bioquímico Phoebus Levene (Foto de
la izquierda) determinó que el DNA estaba formado por 4 tipos
distintos de nucleótidos. Cada nucleótido estaba formado
por desoxirribosa, fosfato y una base nitrogenada (A, C, T o G).
En 1949, el bioquímico Erwin Chargaff (Foto de la derecha)
analizó el contenido molar de las bases de DNA procedente de diversos
organismos y descubrió que en todos los casos [A]=[T] y
que [G]=[C], o lo que es lo mismo, [A+G]=[T+C] ([purinas]=[pirimidinas]).
Esta es la llamada ley de Chargaff.
En
los años 20, el bioquímico Phoebus Levene (Foto de
la izquierda) determinó que el DNA estaba formado por 4 tipos
distintos de nucleótidos. Cada nucleótido estaba formado
por desoxirribosa, fosfato y una base nitrogenada (A, C, T o G).
En 1949, el bioquímico Erwin Chargaff (Foto de la derecha)
analizó el contenido molar de las bases de DNA procedente de diversos
organismos y descubrió que en todos los casos [A]=[T] y
que [G]=[C], o lo que es lo mismo, [A+G]=[T+C] ([purinas]=[pirimidinas]).
Esta es la llamada ley de Chargaff.






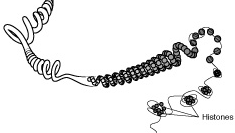
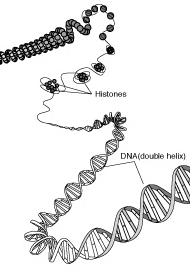
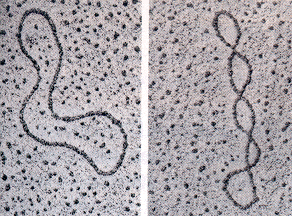 En
procariotas, así como en las mitocondrias y cloroplastos, el DNA se
presenta en forma circular, en la que la doble hélice se cierra por sus
extremos. Este DNA circular puede presentar diversos grados de superenrrollamiento
(Figura de la derecha).
En
procariotas, así como en las mitocondrias y cloroplastos, el DNA se
presenta en forma circular, en la que la doble hélice se cierra por sus
extremos. Este DNA circular puede presentar diversos grados de superenrrollamiento
(Figura de la derecha).